Меристемы
Меристе́мы (с др.-греч. — «μεριστός» — делимый), или образовательные ткани, или меристематические ткани, — обобщающее название для тканей растений, состоящих из интенсивно делящихся и сохраняющих физиологическую активность на протяжении всей жизни клеток, обеспечивающих непрерывное нарастание массы растения и предоставляющих материал для образования различных специализированных тканей (проводящих, механических и т. п.).

Содержание
Особенности клеток меристем
Эумеристемы, дающие начало всем тканям растения, кроме проводящих, состоят из изодиаметрических, в очертании многоугольных клеток. Каждая клетка эумеристемы имеет крупное ядро, занимающее около половины объёма клетки, а ядерная оболочка имеет много пор. В цитоплазме много диффузно расположенных рибосом. Имеются пропластиды, митохондрии и диктиосомы. Вакуоли немногочисленные и мелкие. Соседние клетки соединены более или менее диффузно расположенными плазмодесмами.
Клетки меристем, образующих проводящие ткани (камбий и прокамбий), прозенхимные, снабжены крупными вакуолями.
Клеткам меристем свойственно неравное (дифференцированное) деление. Клетка делится митозом на 2: одна остаётся клеткой меристемы, а другая, поделившись один или несколько раз, приступает к дифференцировке.
Митотическая активность клеток меристемы различна. Наиболее активные в этом отношении клетки — инициальные клетки, дающие начало производным клеткам. Производные могут отличаться от инициальных формой, размерами, количеством вакуолей.
Увеличение общего объёма меристемы может быть следствием только инициальных клеток (так, например, нарастают талломы некоторых бурых водорослей), либо чередованием делений инициальной клетки и производных, причём большее количество делений приходится на производные клетки. Такая периодичность свойственна конусам нарастания большинства высших споровых. У семенных же инициальная клетка и производные морфологически неразличимы.
Меристемы заново образуются из других клеток в повреждённых тканях и отвечают за заживление ран.
Классификация меристем и их производных
Морфологическая классификация
Морфологическая классификация основана на ориентации перегородок в делящихся клетках. Выделяется три морфологических типа меристем:
- Пластинчатые меристемы однослойные, клетки, таблитчатые на поперечном срезе, делятся антиклинально (перпендикулярно плоскости органа). Участвует в образовании эпидермы.
- Колончатые, или стержневые меристемы состоят из кубических или призматических клеток, располагающихся продольными рядами. У некоторых растений такие меристемы образуют сердцевину стебля.
- Массивные меристемы состоят из многоугольных в очертании клеток, делящихся в разных направлениях и обеспечивающих более или менее равномерное увеличение объёма ткани. Из массивных меристем развивается, например, спорогенная ткань спорангия.
Онтогенетическая классификация
Все меристемы, функционирующие на протяжении всей жизни растения, можно подразделить на общие и специальные. К общим меристемам относится меристема зародыша, из которой развиваются апикальные (верхушечные) меристемы, а они, в свою очередь, дают начало третьей группе общих меристем — интеркалярным.
От апикальных меристем берут начало:
- протодерма, образующая эпидерму;
- основная меристема, дающая начало системе тканей основной паренхимы;
- прокамбий.
Клетки этих трёх тканей более вакуолизированы, чем апикальные меристемы, и обладают ограниченной способностью к делению, поэтому часто их называют полумеристемами.
Часть клеток прокамбия даёт начало первичным проводящим тканям — первичным ксилеме и флоэме; из других его клеток развивается камбий, который, в свою очередь, образует вторичные проводящие ткани — вторичные ксилему и флоэму. Вторичные проводящие ткани образует и т. н. добавочный камбий, развивающийся не из прокамбия, а из постоянных тканей, живые клетки которых при этом дедифференцируются, то есть возвращаются из специализированного состояния к состоянию, близкому к эмбриональному, и вновь приобретают способность к делению.
Таким же образом в растении возникают и раневые меристемы, восстанавливающие повреждённые участки растения. На возможности новообразования меристем основано вегетативное размножение растений.
Кроме добавочного камбия, постоянные ткани также участвуют в образовании феллогена (пробкового камбия). Делясь периклинально (то есть параллельно поверхности органа), клетки феллогена отделяют наружу будущие клетки пробки (феллемы), а внутрь — феллодерму.
Топографическая классификация
В соответствии с расположением, помимо апикальных меристем, локализованных на кончиках осевых органов — стебля и корня, выделяют меристемы:
- латеральные (боковые) меристемы расположены внутри осевых органов и осуществляют их утолщение. К латеральным меристемам относят камбий, добавочный камбий и феллоген. Они являются вторичными меристемами, потому что они ответственны за вторичный рост или увеличение обхвата и толщины ствола.
- краевая (маргинальная) меристема даёт начало листовой пластинке.
- интеркалярные меристемы осуществляют вставочный рост. За счёт их деятельности удлиняются междоузлия на ранних этапах развития побега, развиваются черешки листьев. Принципиально они отличаются от прочих меристем тем, что, помимо недифференцированных клеток, в них существуют и клетки на разных стадиях дифференциации. Поэтому корректнее здесь говорить не об интеркалярных меристемах, а о зонах интеркалярного (вставочного) роста.
Все эти меристемы, кроме феллогена, являются производными апикальных меристем.
Генетическая классификация
Согласно генетической классификации выделяют первичные и вторичные меристемы. Первичные меристемы связаны с меристемами зародыша и апикальными меристемами, а вторичные развиваются позднее, когда начинается утолщение осевых органов и заканчивается верхушечный рост. Однако такое разделение условно, так как, например, камбий закладывается до окончания верхушечного роста.
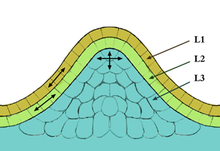
Апикальная меристема
Развивающийся зародыш любого растения состоит из эумеристемы. По мере формирования зачатков вегетативных органов у зародыша эумеристема остаётся только в двух местах: в верхушке зачаточного побега (будущий конус нарастания). Эти меристемы и называют апикальными меристемами (от лат. apex — вершина). Они осуществляют рост побегов и корней в длину и дают начало большинству других меристем.
Когда растение вступает в фазу цветения, верхушечная меристема стебля преобразуется в меристему соцветия. От неё берёт начало флоральная меристема, образующая структуры цветка: лепестки, чашелистики, плодолистики и тычинки.
- Лотова Л. И. Ботаника: Морфология и анатомия высших растений. — Изд. 4-е, доп.. — М.: Книжный дом «ЛИБРОКОМ», 2010. — 512 с. — ISBN 978-5-397-01047-4.
- Эсау К. Анатомия растений, пер. с англ.. — М., 1969.
- Lehrbuch der Botanik für Hochschulen, 30 Aufl.. — Jena, 1971.